Moving forward: insights and applications of moving‐habitat models for climate change ecology
Published in Journal of Ecology, 2017.
Harsch MA, Phillips A, Zhou Y, Leung MR, Rinnan DS and Kot M (2017). "Moving forward: insights and applications of moving‐habitat models for climate change ecology." Journal of Ecology, 105(5), 1169-1181. https://doi.org/10.1111/1365-2745.12724 Download here
Summary:
Predicting and managing species’ responses to climate change is one of the most significant challenges of our time. Tools are needed to address problems associated with novel climatic conditions, biotic interactions and greater climate velocities.
We present a spatially explicit moving‐habitat model (MHM) and demonstrate its versatility in tackling critical questions in climate change research, including dispersal in multiple spatial dimensions, population stage structure, interspecific interactions, asymmetric range shifts, Allee effects and the presence of infectious diseases. The model utilizes integrodifference equations to track changes in population density over time in a habitat that is moving. The model is quite flexible and can accommodate variation in demography, dispersal patterns, biotic interaction and stochasticity in the velocity of climate change.
The methods provide a general mechanistic understanding of the underlying ecological processes that drive a system. Field data can be readily incorporated into the model to give insight into specific populations of interest and inform management decisions.
Synthesis. Moving‐habitat models unite ecological theory, data‐centred modelling and conservation decision support under a single framework. Their ability to generate testable hypotheses, incorporate data and inform best management practices proves that these models provide a valuable framework for climate change biologists.
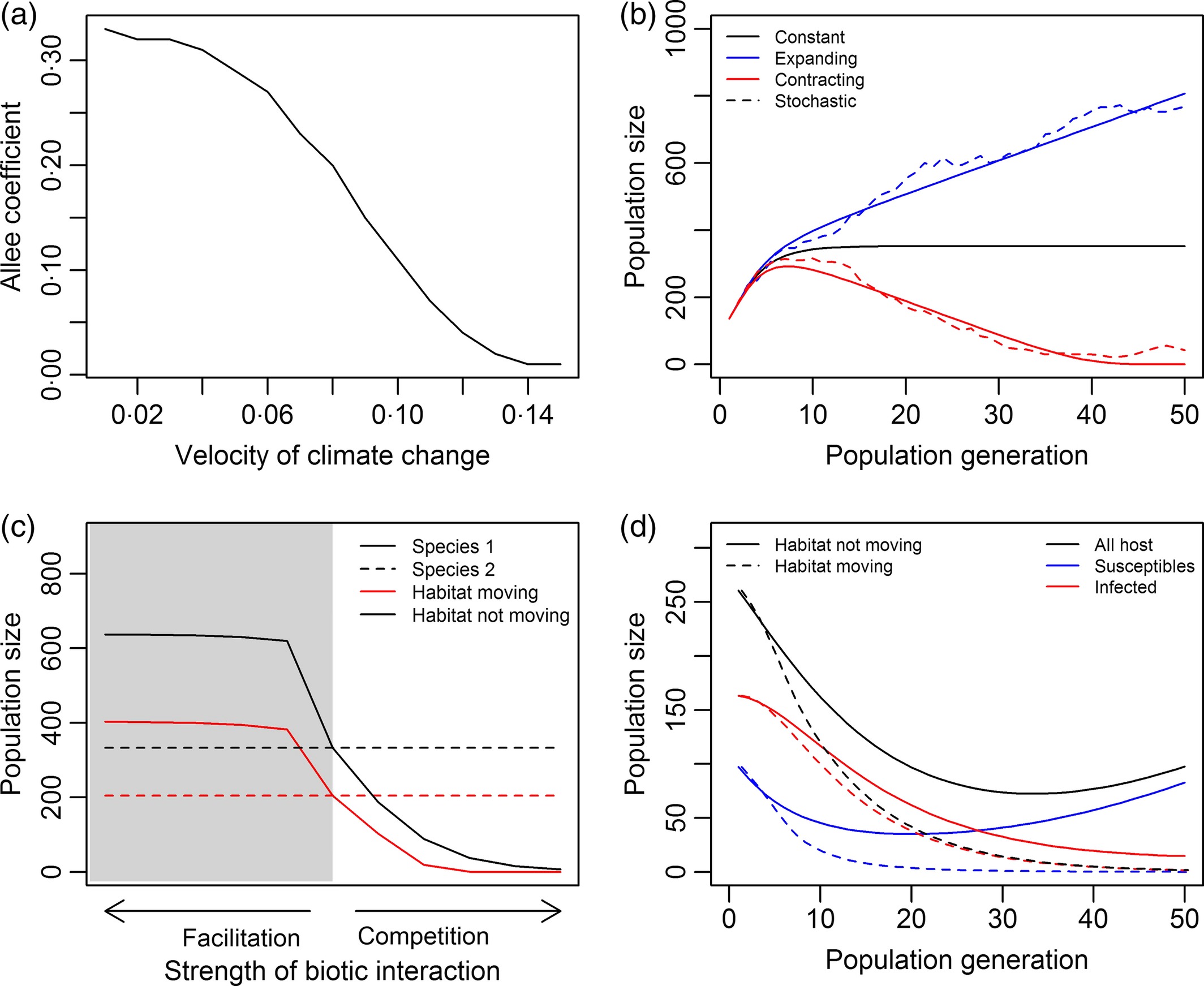
Figure 5: (a) The maximum Allee coefficient for a given velocity of climate change resulting in a population size greater than 0 after 50 time steps. In this case study, the mean dispersal distance is slightly greater than the velocity of climate change ($c$ + 0.8 km). (b) Effect of asymmetric shift rates on population size. Solid lines indicate results when $c$ (0.5 in this case study) is deterministic and dashed lines when c is stochastic. Line colour indicates results when habitat size does not change (black line), is expanding (blue line) or contracting (red line). The mean dispersal distance is set to 1 km. (c) Change in total population size of a species after 50 times steps as the strength of the biotic interaction (facilitation or competition) by an interacting species increases on a stationary (black lines) and a moving habitat (red lines). In this example, species 2 (dashed lines) affects species 1 (solid lines) but species 1 has no effect on species 2. Shaded section of graph represent when species 2 facilitates species 1. We set $c$ to 1 and the mean dispersal distance for both species to 1·5 km. (d) Change in total population size (black lines), the size of the subpopulation infected (red lines), and the size of the subpopulation susceptible but not yet infected (blue lines), both when the habitat is not moving (solid lines) and when the habitat is moving (dashed lines). We set $c$ to 1, the mean dispersal distance to 0.75 km for the host species and 1 km for the infectious agent, a maximum reproduction rate of 20 and the population density at which half the maximum are produced at 100, a mortality of 10% for infected individuals, and a half‐saturation constant of infection of 150. Initially, 60% of the host population are infected. In all case studies, the habitat length is set to 5 km, and we specify a Gaussian dispersal kernel, a carrying capacity of 100, and a net reproductive rate of 2.